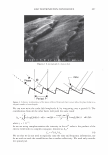
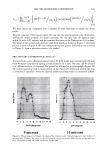
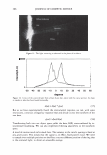
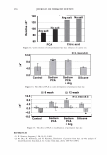
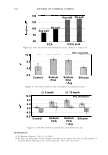
2006 TRI/PRINCETON CONFERENCE 365 120 110- 100- E 90- � � 80- :i.... Q) 70- - $ Q) E 60- «1 :a 50- 40- 30 Figure 7. Diameters data for unmedullated (white box) and medullated hair (gray box). Box plots were obtained from 40 hair fiber measurements from each sample. ca 500- Cl.. � 400- .c - � C) C: Q) 300- :i.... - Cl) C) C: 200- :2 «1 Q) :i.... 100- cc Figure 8. Breaking strength data for unmedullated (white box) and medullated hair (gray box). Box plots were obtained from 40 hair fiber measurements from each sample. 70�------------------, 65 '#- c: 60 0 � 55 C) 5 50 a 45- C) C: :2 40- «1 � 35- .0 30...____________________. Figure 9. Breaking elongation data for unmedullated (white box) and medullated hair (gray box). Box plots were obtained from 40 hair fiber measurements from each sample.

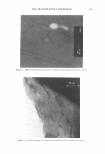
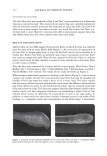
366 JOURNAL OF COSMETIC SCIENCE 9000 8000 a. $ � - 7000 ::::J 6000 5000 1 4000 3000 Figure 10. Modulus data for unmedullated (white box) and medullated hair (gray box). Box plots were obtained from 40 hair fiber measurements from each sample. terizes the yield region. Once the post-yield region is reached, the remaining 70% of the a-helices unfold irreversibly by a compulsory breakdown of the disulphide bonds and they constitute the Y zone. The series zone concept was modified (26) with an extended two phase model to explain the increase of stress in the post-yield region. This model suggests that the increased stress is produced by globular matrix proteins jamming the microfibrils as the fiber is extended into the post-yield region. In this model, water molecules are supposed to be ejected from the matrix at high stress levels, which leads to matrix protein compression between the microfibrils (14). In Chapman's model (27) matrix proteins are supposed to be covalently bonded to fundamental repeat units aligned along the microfibril. The stress-strain curve of the fiber reflects the permanent interaction between microfibril and matrix. Considering TEM observations for medulla, there are several reasons for expecting different mechanical behavior for medullated hairs. First, the microfibrils are randomly distributed through a 20% of the diameter of the fiber. Thus, most of them are not aligned in the axial direction and this could affect the yield region. According to Clement et al. (22), the structures in medulla are cross-linked by peptide bonds from citrulline residues. They might not have the same tensile resistance as the disulfide bonds of the matrix. But when the data is displayed in box plots (Figures 7-10) it is noted that average values are the similar for medullated and unmedullated hair in most cases but that medullated hair shows broader distributions. Considering the data of Figure 7, unmedullated hair has about 5 7 µm of diameter or "pure cortex". On the other hand, medullated hair has about 78 µm of diameter but has a porous inner structure of about 15 µm. Deducting the diameter of medulla, medullated hair has about 63 µm of diameter of "pure cortex". This means that both types of hair should show similar mechanical behaviors. The higher variance in stress-strain curves of medullated hair could be caused by the difference between the fiber diameter minus the medulla diameter. Also, the cavities in the medulla structure as mechanical defects generating cracks which lead to ultimate failure. CONCLUSIONS TEM micrographs allowed the observation of cortical cells disposed randomly in the medulla. The main evidence was the observation of microfibrils.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)